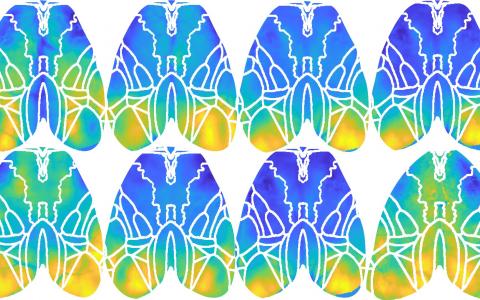
Decision-making using uncertain sensory evidence and temporal expectation
15 April 2021
By April Cashin-Garbutt
How does sensory evidence get processed by the brain and culminate in an action? This question has been challenging to answer, as a lot of decision-making processes are correlated with one another and embedded in ongoing brain activity. While previous research in neuroscience has focused on studying decision-making in human and non-human primates, large efforts have been made recently to study decision-making in mice.
“Mouse models are very powerful in neuroscience. While mice are not capable of making decisions that are as complex as those made by humans, mouse models offer the chance to study the cellular and circuit mechanisms underlying decision-making in much greater detail. ” said Ivana Orsolic, Research Fellow in the Mrsic-Flogel Lab.
Researchers at the Sainsbury Wellcome Centre explored decision-making in mice using a novel behavioural framework and widefield calcium imaging across dorsal cortex to elucidate how sensory evidence gets transformed across multiple brain areas simultaneously. In addition, the team studied how the process of transformation of sensory evidence is influenced by temporal expectation.
Novel behavioural framework
To understand the interaction of sensory evidence and temporal expectation in decision-making, Ivana and team designed a new behavioural task for mice.
“Our task design was inspired by gold standards set in primate decision-making, mainly parametrisation of the stimulus and careful motor control. A twist in the task design allowed us to separate processes of choice execution from processes of ongoing decision-making and prolonged the window of ongoing decision-making onto timescales typically not studied. This allowed us to ask how the mouse’s prior knowledge of the task structure, including temporal expectation, interacts with current sensory evidence,” commented Ivana.
While the mice were voluntarily stationary, they were shown a complex, changing visual stimulus under experimenter’s control and they had to detect when the changes in the stimulus were sustained. The mice didn’t know exactly when the change would happen or the amplitude of the change, so they had to pay attention to small changes throughout. In addition to the requirement to observe the sensory evidence and be patient, the mice were encouraged to report as soon as they believed that the change had occurred. This was achieved by limiting the response window when the reward was available.
Separating choice execution from ongoing decision-making
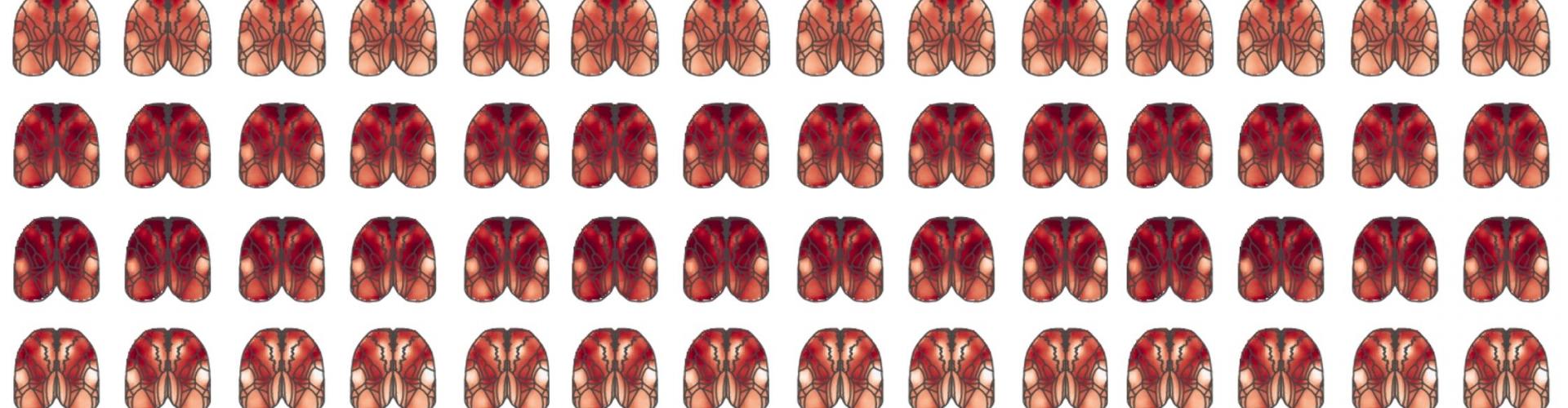
Researchers characterised the neuronal activity in large areas of neocortex in the period of ongoing decision-making prior to choice execution. As expected, while the mice observed the visual stimulus change, activity of the primary visual areas was modulated earliest, followed by higher visual areas, culminating in secondary motor cortex. While visual areas were also engaged in naïve animals, this engagement of secondary motor cortex by the sensory stimulus was present only in expert animals.
Video shows how sensory inputs propagate in the cortical activity of mice performing the task (left) and untrained mice (right) viewing the same stimulus.
The team also looked to see how long a given stimulus affects the relevant brain areas. In visual areas, they saw that this history was very short – the primary visual areas would activate and inactivate very quickly. This again was to be expected as the primary visual areas represent the change in stimulus.
In contrast, the secondary motor cortex was modulated by the stimulus on long timescales and this occurred seconds before the animal would report its choice. The way visual evidence modulates the secondary motor cortex was also very interesting. Unlike in visual areas – where any short-term increases or decreases in the speed of the stimulus affect the activity – in the secondary motor cortex, only increases in the speed (which signal that change is likely to occur and mice should respond) affected activity. This shows that secondary motor cortex is more selectively engaged by the visual stimulus and this engagement is sustained, compared to visual areas.
Furthermore, the researchers found that secondary motor cortex was modulated by temporal expectation. The selective representation was influenced by prior expectation of the mouse over whether change was likely to occur.
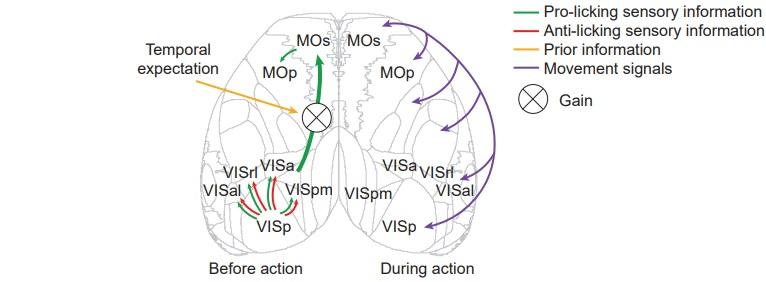
Figure shows the patterns of cortical activation during sensorimotor transformation in the change detection task. Left: activity of dorsal cortex during processing of sensory evidence. While sensory evidence modulates visual areas bidirectionally, motor cortical areas are selectively modulated by sensory information that indicates that the change may have occurred. This modulation occurs only in trained animals and outlasts the stimulus presentation itself. Right: around the time of action execution, cortical activity is broadly modulated by animals' motor output, contrasting with the more specific cortical activity patterns during deliberation.
Recruitment of secondary motor cortex
The researchers were surprised to find that recruitment of secondary motor cortex by sensory evidence in this task was modulated by temporal expectation.
“We were very careful about monitoring body movements of the mouse during the task using videography. Even when accounting for small facial movements that a mouse would make, we confirmed this selective engagement of secondary motor cortex by sensory evidence. We were very surprised to find that this relationship was modulated by temporal expectation of the mouse. This is an exciting finding as it suggests emerging motor plan in secondary motor cortex that tracks both sensory evidence and expectation from the onset of the trial, long ahead of the choice execution, or that secondary motor cortex represents more abstract aspects of decision-making, like the mouse’s belief about the state of the stimulus,” said Ivana.
In future work, Ivana and team hope to dissect these processes in secondary motor cortex further to see if they can separate belief from temporal expectation and motor preparation. They plan to design a new task variant to manipulate the behaviour and hope to find a way of tracking what the animal currently believes and relate that belief to both the behaviour and the underlying neural activity.
Read the paper in Neuron: Mesoscale cortical dynamics reflect the interaction of sensory evidence and temporal expectation during perceptual decision-making